
餐廚廢水厭氧消化沼氣生物提純氣體循環(huán)技術(shù)
近年來(lái),餐廚垃圾的產(chǎn)量急劇增加,據(jù)統(tǒng)計(jì),中國(guó)每年產(chǎn)生的餐廚垃圾超過(guò)6000×104 t。在餐廚垃圾的處理過(guò)程中會(huì)產(chǎn)生大量的餐廚廢水,而餐廚廢水的無(wú)害化和資源化處理受到廣泛關(guān)注。厭氧消化技術(shù)可以將餐廚廢水中的有機(jī)質(zhì)轉(zhuǎn)化為沼氣,實(shí)現(xiàn)能源的回收與利用。然而,厭氧消化產(chǎn)生的沼氣中含有40%~60%的不可燃?xì)怏wCO2,降低了沼氣的熱值。有研究表明,提純后的沼氣(CH4>90%)可用作車(chē)輛燃料或作為天然氣替代能源。目前,沼氣提純方法可以分為物理法、化學(xué)法和生物法等。沼氣生物提純作為一種新型處理技術(shù)越來(lái)越受到重視,該技術(shù)利用嗜氫產(chǎn)甲烷菌的新陳代謝作用將CO2和H2轉(zhuǎn)化為CH4,而不是將CO2從體系中分離或吸收,可減少CO2排放,故具有低能耗、無(wú)二次污染等特點(diǎn)。
根據(jù)供氫方式的不同,沼氣生物提純技術(shù)可分為原位提純和異位提純。原位沼氣提純中H2直接注入?yún)捬跸w系中,在微生物作用下H2和CO2轉(zhuǎn)化為CH4,裝置簡(jiǎn)單易操作,但氫分壓的增加及混合剪切力的增強(qiáng)可能會(huì)降低產(chǎn)甲烷菌的性能,導(dǎo)致有機(jī)酸積累,降低反應(yīng)效率。異位生物提純中厭氧消化和沼氣提純?cè)?個(gè)獨(dú)立體系內(nèi)進(jìn)行,厭氧消化產(chǎn)生的沼氣通入提純體系內(nèi),嗜氫產(chǎn)甲烷菌在H2的作用下提純沼氣,具有較高穩(wěn)定性和靈活性。然而,H2的氣液傳質(zhì)效率成為沼氣提純的主要限制因素。如何提高H2的溶解度是解決低H2傳質(zhì)速率的關(guān)鍵。納米零價(jià)鐵(nanozero-valentiron,nZVI)在厭氧條件下可以與水反應(yīng)生成H2(式(1)),該過(guò)程中H2從容器底部析出,可以加大氣液接觸面積。DONG等的研究表明,nZVI溶解產(chǎn)生H2可以提高CO2的甲烷化。此外,氣體傳遞系數(shù)的提高可以提升H2氣-液傳質(zhì)速率(式(2))。有研究表明,氣體循環(huán)可有效增加氣體傳遞系數(shù),提高H2利用率和CH4含量。KARIM等利用氣體循環(huán)提高了牛糞厭氧體系中CH4的產(chǎn)量。ZHAO等在氣體循環(huán)條件下對(duì)污泥進(jìn)行半連續(xù)厭氧消化處理,與無(wú)氣體循環(huán)相比,CH4含量提高了14%~15%。但關(guān)于nZVI供氫的異位生物提純與氣體循環(huán)相結(jié)合的研究報(bào)道還較少。
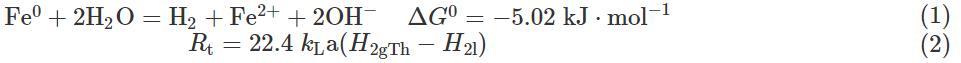
式中:Rt表示H2傳質(zhì)速率,mol·(L·h)-1;kLa表示線(xiàn)性氣體傳遞系數(shù),h-1;H2gTh表示氣相中H2濃度,mol·L-1;H2l表示液相中H2濃度,mol·L-1。
本研究通過(guò)厭氧消化處理餐廚廢水,采用異位生物提純方法提高CH4含量。在實(shí)驗(yàn)過(guò)程中,利用nZVI在厭氧體系下析氫為沼氣提純體系提供H2,同時(shí)設(shè)計(jì)單向式、往復(fù)式、循環(huán)式3種氣體循環(huán)方式,分析了不同氣體循環(huán)方式下沼氣異位生物提純效果,以期獲得最佳的運(yùn)行方式。
1、材料與方法
1.1 實(shí)驗(yàn)材料
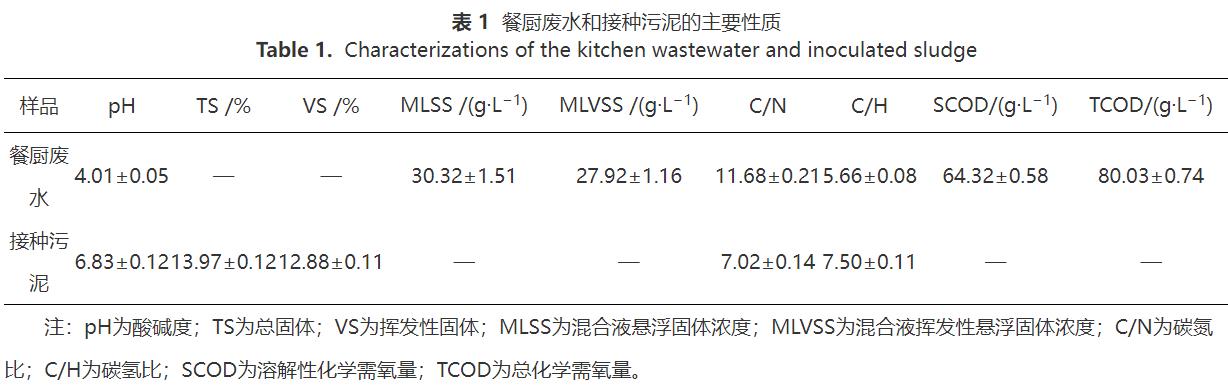
厭氧消化的底物為餐廚廢水,取自無(wú)錫市某餐廚垃圾處理廠。厭氧顆粒污泥取自無(wú)錫市某食品廠的厭氧消化池。原始的餐廚廢水和接種污泥的性質(zhì)如表1所示。nZVI(直徑50nm,純度99.9%)購(gòu)自寧波金雷納米材料科技有限公司。
1.2 實(shí)驗(yàn)裝置
本研究設(shè)置了單向式、往復(fù)式和循環(huán)式3種實(shí)驗(yàn)。每種方式均采用異位生物提純法,由厭氧消化體系(圖1,A1、A2、A3)和沼氣提純體系(圖1,U1、U2、U3)2個(gè)體系構(gòu)成。其中,前一體系用于餐廚廢水厭氧消化處理,產(chǎn)生沼氣;后一體系用于沼氣生物提純,nZVI提供H2。在前期研究基礎(chǔ)上,將收集的餐廚廢水經(jīng)離心去除固體物質(zhì)后稀釋10倍使用,厭氧消化體系中底物和接種物按COD底物:VS接種物(質(zhì)量比)為1.2∶1在500mL血清瓶?jī)?nèi)接種。將nZVI和厭氧顆粒污泥投加到沼氣生物提純體系中,根據(jù)文獻(xiàn)報(bào)道和前期實(shí)驗(yàn),選擇nZVI的質(zhì)量濃度為6g·L-1,該體系不添加其他底物,實(shí)驗(yàn)設(shè)計(jì)參數(shù)如表2所示。實(shí)驗(yàn)前向所有反應(yīng)瓶?jī)?nèi)通5min氮?dú)庖跃S持厭氧環(huán)境,采用2mol·L-1 NaOH或HCl將厭氧-沼氣提純反應(yīng)瓶?jī)?nèi)初始pH調(diào)至7.00,在(36±1)℃下反應(yīng),反應(yīng)過(guò)程中對(duì)厭氧消化體系采用間歇式攪拌,間隔時(shí)間為2h,攪拌轉(zhuǎn)速為60r·min-1。實(shí)驗(yàn)的前3d為厭氧消化產(chǎn)沼氣,從第4天開(kāi)始啟動(dòng)沼氣循環(huán)操作,每天循環(huán)2次,每次循環(huán)持續(xù)1h,具體操作方式如下文所述。
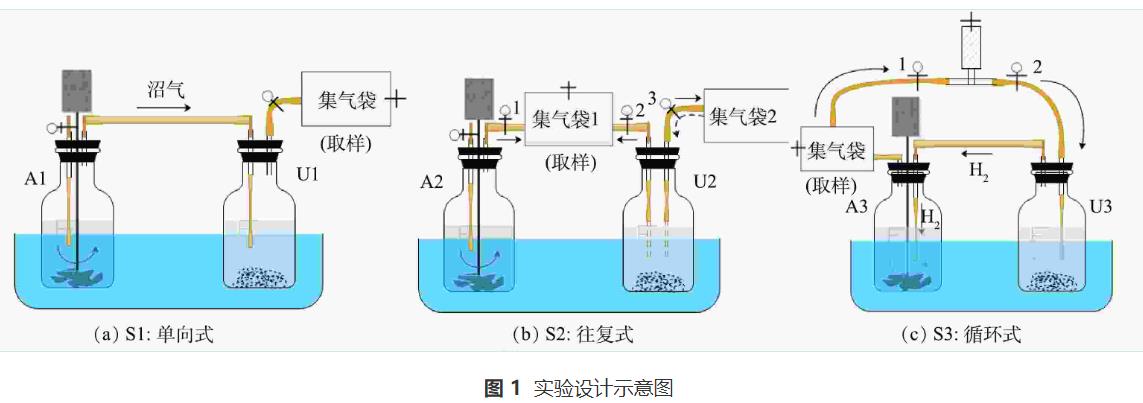
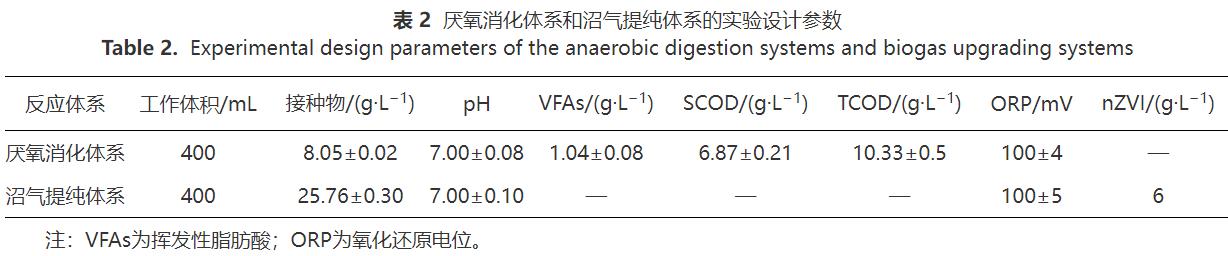
實(shí)驗(yàn)S1為單向式。在A1內(nèi)進(jìn)行餐廚廢水厭氧消化處理,產(chǎn)生的沼氣(含CO2)通入U(xiǎn)1體系,在U1內(nèi)利用nZVI供氫實(shí)現(xiàn)沼氣生物提純。
實(shí)驗(yàn)S2為往復(fù)式。在最初3d內(nèi),打開(kāi)閥門(mén)1和2,關(guān)閉閥門(mén)3,在集氣袋1內(nèi)收集A2產(chǎn)生的沼氣(含CO2)以及U2中nZVI產(chǎn)生的H2。第4天啟動(dòng)氣體循環(huán),做氣體循環(huán)處理時(shí),關(guān)閉閥門(mén)1,打開(kāi)閥門(mén)2和3,以往復(fù)方式循環(huán)集氣袋1中收集的氣體。往復(fù)循環(huán)時(shí)先將U2反應(yīng)瓶?jī)?nèi)混合物搖勻,再將U2反應(yīng)瓶?jī)A斜,使閥門(mén)2連接的氣管進(jìn)入到U2反應(yīng)瓶液面以下(圖1-U2中虛線(xiàn)氣管),集氣袋1內(nèi)氣體全部通入U(xiǎn)2,經(jīng)微生物利用后進(jìn)入集氣袋2。然后將閥門(mén)3連接的氣管進(jìn)入到U2反應(yīng)瓶液面以下(圖1-U2中虛線(xiàn)氣管),集氣袋2內(nèi)氣體全部通入U(xiǎn)2,經(jīng)微生物利用后進(jìn)入氣袋1。重復(fù)上述操作持續(xù)1h,每天2次。不做氣體循環(huán)處理時(shí),打開(kāi)閥門(mén)1和2,關(guān)閉閥門(mén)3,在集氣袋1內(nèi)收集A2和U2產(chǎn)生的氣體。
實(shí)驗(yàn)S3為循環(huán)式。在最初的3d內(nèi),關(guān)閉閥門(mén)1和2,U3中nZVI產(chǎn)生的H2先進(jìn)入A3體系,經(jīng)微生物利用后與A3中產(chǎn)生的沼氣混合進(jìn)入集氣袋。第4天啟動(dòng)氣體循環(huán),打開(kāi)閥門(mén)1和2,將集氣袋內(nèi)的氣體依次通入U(xiǎn)3、A3,形成U3-A3-集氣袋-U3閉環(huán)沼氣循環(huán),重復(fù)上述操作持續(xù)1h,每天2次。
1.3 分析方法
TS、VS、MLSS、MLVSS、COD測(cè)定根據(jù)《國(guó)家水和廢水監(jiān)測(cè)分析方法》。pH采用pH計(jì)測(cè)定,ORP采用氧化還原電位儀測(cè)定,富氫值(HE)采用富氫電極測(cè)定,C、N、H采用元素分析儀測(cè)定。VFAs濃度采用氣相色譜儀(日本島津GC-2010PLUS)測(cè)定,以高純氮為載氣,檢測(cè)器采用氫火焰離子化檢測(cè)器(FID),溫度設(shè)置為250℃;分析柱為peg-20m色譜柱,柱溫設(shè)置為70℃;進(jìn)樣口溫度為250℃。氣體(H2、CH4和CO2)采用氣相色譜儀(GC-2014,日本島津公司)測(cè)定,熱導(dǎo)檢測(cè)器(TCD)及TDX-1色譜柱,柱溫,進(jìn)樣口溫度和檢測(cè)器溫度分別為100、180和180℃,載氣為氬氣。根據(jù)GOEL等方法測(cè)定蛋白酶和脫氫酶的活性,按照DU等方法測(cè)定乙酸激酶活性,通過(guò)分光光度法測(cè)定輔酶F420活性。
1.4 微生物群落分析
各反應(yīng)體系結(jié)束后采集污泥樣品,使用PowerSoil試劑盒從樣品中提取DNA,再進(jìn)行聚合酶鏈反應(yīng)(PCR)擴(kuò)增。細(xì)菌的引物為341F(5′-CCTACGGGGNGGCWGCAG-3′)和805R(5′-GACTAChVGGTATCTACC-3′)。古菌第1輪引物為340F(5′-CCCTayGGGYGCASCAG-3′)和1000R(5′-GGCCATGCACYWCYTCTC-3′),第二輪引物為349F(5′-GYGCASCAGKCGMGAAW-3′)和806R(5′-GGACTACVSGGGTATCTAAT-3′),測(cè)序平臺(tái)為IlluminaMiSeq。
2、結(jié)果與討論
2.1 不同氣體循環(huán)方式對(duì)氣體含量變化的影響
如圖2(a)所示,前3d沒(méi)有啟動(dòng)氣體循環(huán),S1、S2、S3的CH4含量均呈現(xiàn)上升的趨勢(shì)。啟動(dòng)循環(huán)后,與S1相比,S2和S3的CH4含量在4~10d內(nèi)迅速增加。S1、S2、S3的CH4含量在第10天分別達(dá)到76.46%、87.15%、92.63%,在提純過(guò)程中的最大CH4含量分別為81.69%、88.12%、93.87%。有研究表明,未經(jīng)提純的沼氣中CH4含量約為60%。本實(shí)驗(yàn)中經(jīng)異位生物提純后CH4含量顯著提高(P<0.05),與單向式相比,往復(fù)式和循環(huán)式氣體循環(huán)后CH4含量分別提高了6.43%和12.18%。
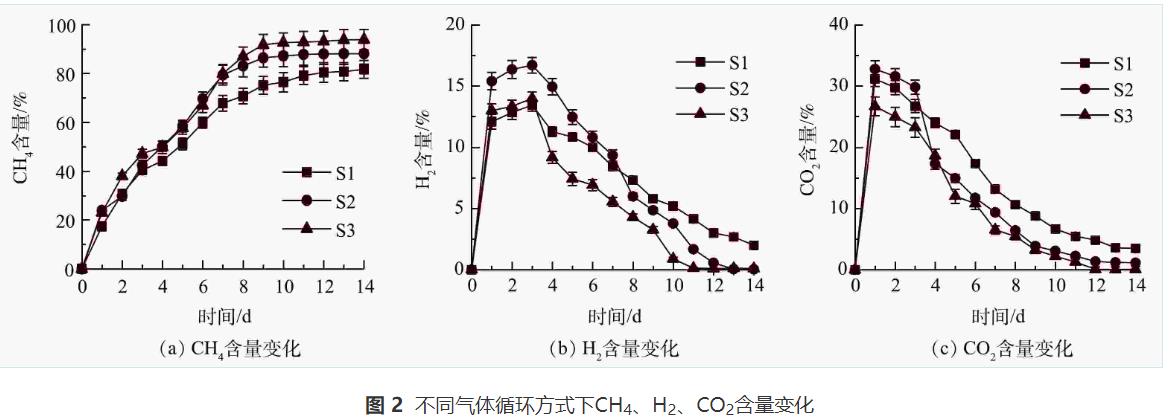
由圖2(b)可以看出,由于nZVI腐蝕析氫和沼氣的產(chǎn)生,S1、S2和S3的H2含量在第3天達(dá)到最大值,即13.46%、16.70%、14.00%,且S1和S3中的H2含量顯著低于S2(P<0.05)。這主要是因?yàn)椋篠1和S3體系首先啟動(dòng)了異位生物提純,H2的產(chǎn)生與消耗同時(shí)進(jìn)行;而S2產(chǎn)生的H2在前3d內(nèi)直接進(jìn)入集氣袋內(nèi)不參與沼氣提純。在啟動(dòng)氣體循環(huán)后,H2被反復(fù)注入體系并被微生物利用,在反應(yīng)結(jié)束時(shí)幾乎檢測(cè)不到H2,CH4含量也隨著H2循環(huán)次數(shù)的增加而提高。此外,H2的再循環(huán)也為厭氧消化體系提供更多的底物,促進(jìn)了有機(jī)物的水解,生物提純體系中nZVI與水產(chǎn)生H2的同時(shí)還生成Fe2+,F(xiàn)e2+可以與S2-結(jié)合,減少氣體循環(huán)過(guò)程中H2S的累積,從而避免H2S對(duì)產(chǎn)甲烷菌的抑制。
圖2(c)為CO2含量的變化情況。與H2變化相似,不同的是啟動(dòng)氣體循環(huán)后,CO2的降解速率高于H2。這主要是由于CO2在水中有較高的溶解性。含有H2和CO2的氣體循環(huán)可增加H2和CO2的溶解,并提高厭氧消化體系內(nèi)氣-液-固相的接觸時(shí)間,這有利于CH4的產(chǎn)生。LATHA等的研究表明,由于CO2的酸化性,間歇式氣體循環(huán)可以促進(jìn)酸堿平衡。在本實(shí)驗(yàn)中,nZVI在水中的溶解會(huì)產(chǎn)生OH-,提高系統(tǒng)的pH,這對(duì)產(chǎn)甲烷微生物產(chǎn)生不利影響,但CO2再循環(huán)可以維持體系pH穩(wěn)定。
圖3為累積沼氣和CH4產(chǎn)量情況。結(jié)果表明,S1、S2、S3反應(yīng)14d后的累積沼氣(CH4)產(chǎn)量分別為70.57(57.65)、96.60(85.12)、167.44(157.18)mL·g-1。與S1和S2相比,S3體系內(nèi)沼氣和甲烷產(chǎn)量均顯著提高(P<0.05)。這與ZHAO等的研究結(jié)果一致。
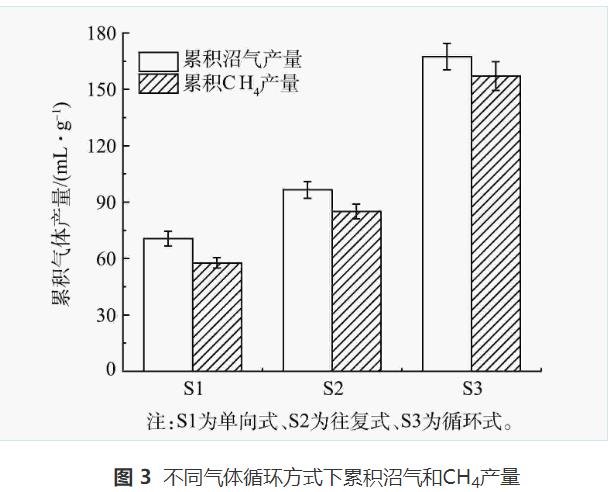
據(jù)報(bào)道,氣體循環(huán)可以提高厭氧消化體系內(nèi)產(chǎn)甲烷菌對(duì)底物的利用率。由于S3中的氣體在厭氧消化和沼氣提純2個(gè)體系中進(jìn)行循環(huán),兩體系的微生物可以將H2和CO2轉(zhuǎn)化為CH4,從而提高S3中的甲烷產(chǎn)量。而S1未進(jìn)行氣體循環(huán),S2的氣體僅在沼氣提純體系中進(jìn)行循環(huán),因此,相對(duì)于S3,S1、S2的H2和CO2利用率更低。
2.2 COD和VFAs的變化情況
沼氣/CH4產(chǎn)量與厭氧反應(yīng)體系中COD和VFAs的降解相關(guān)。如圖4(a)所示,A1、A2和A3的COD值分別從第4天的2856、1776、2032mg·L-1下降到第14天的140、125、115mg·L-1,A2和A3的COD降解速率高于A1。這一結(jié)果表明,氣體循環(huán)有利于促進(jìn)餐廚廢水中有機(jī)物的降解。WEI等認(rèn)為,外源H2不僅可以將CO2轉(zhuǎn)化為CH4,還可以促進(jìn)有機(jī)物降解。氣體循環(huán)可以提高有機(jī)物的產(chǎn)CH4率,這也是S2和S3體系中CH4產(chǎn)量增加的原因之一。
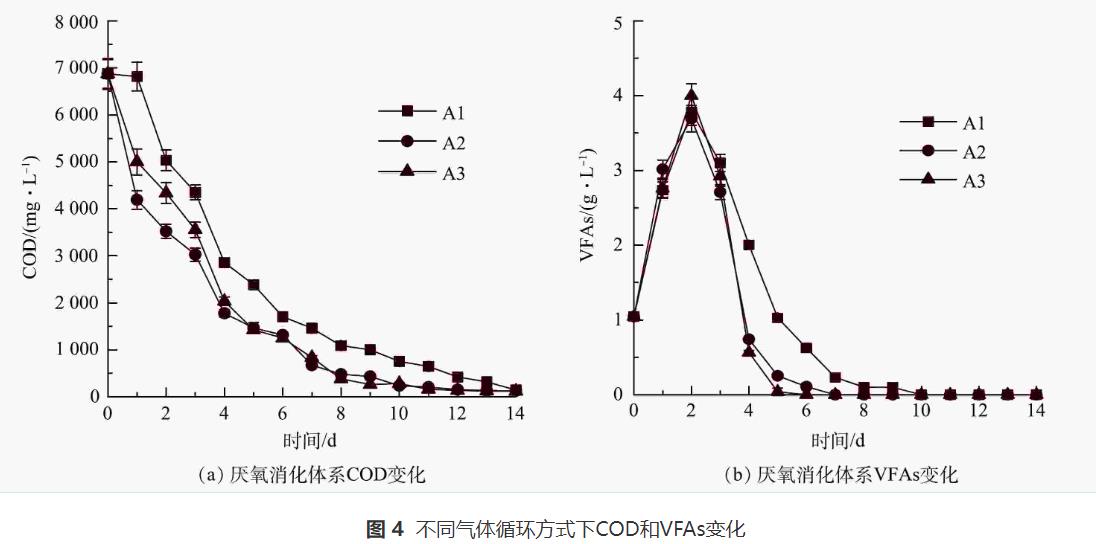
由圖4(b)可以看出,A1、A2、A3的最大VFAs質(zhì)量濃度(第2天)分別為3.78、3.70、3.99g·L-1,此后VFAs逐漸下降直到反應(yīng)結(jié)束。3個(gè)厭氧消化體系均未出現(xiàn)酸化現(xiàn)象,但VFAs的降解率是否與有氣體循環(huán)有關(guān)。在第4天開(kāi)始對(duì)S2和S3啟動(dòng)氣體循環(huán)后,A2和A3中VFAs的降解速率明顯高于A1(P<0.05)。A2和A3系統(tǒng)中的VFAs分別在第6天和第7天完全降解,而A1系統(tǒng)VFAs降解至0的時(shí)間(第10天)相對(duì)滯后。由此可知,氣體循環(huán)可提高厭氧微生物的降解性能。
2.3 pH、ORP、HE的變化情況
由圖5(a)可看出,A1、A2、A3中的pH在厭氧消化開(kāi)始時(shí)由7.00迅速下降到6.75、6.79、6.76,然后上升直至反應(yīng)結(jié)束。這是由于厭氧消化過(guò)程中快速水解和酸化導(dǎo)致VFAs積累,隨著VFAs的消耗,pH逐漸升高。整個(gè)過(guò)程中A1、A2、A3的最高pH分別為7.46、7.39、7.35。與A1相比,由于氣體循環(huán),A2和A3在穩(wěn)定階段更容易維持中性pH。這種現(xiàn)象在沼氣提純體系中較明顯。在U2和U3中由于nZVI溶于水產(chǎn)生OH-,pH在前3d逐漸升高(圖5(b)),這對(duì)產(chǎn)甲烷菌活性和nZVI釋氫均有不利影響。啟動(dòng)氣體循環(huán)后,pH逐漸恢復(fù)到產(chǎn)甲烷菌的最適范圍(6.5~7.5)。LI等認(rèn)為,通過(guò)氣體循環(huán)將CO2多次溶解到沼氣提純液相中,可中和nZVI產(chǎn)生的OH-以維持體系pH穩(wěn)定,為產(chǎn)甲烷菌提供最佳環(huán)境。在U1體系中,從第1天開(kāi)始,pH沒(méi)有顯著提高(P>0.05)。這是由于沼氣異位生物提純過(guò)程中來(lái)自A1的CO2不斷進(jìn)入U(xiǎn)1,因而中和U1中nZVI產(chǎn)生的OH-。
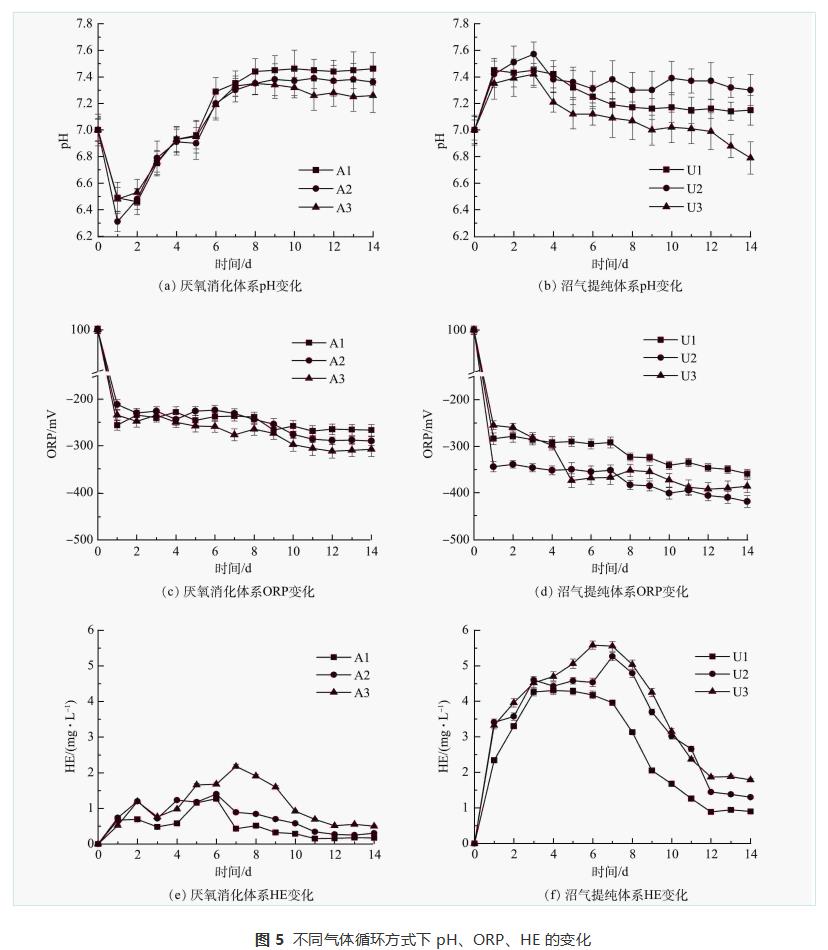
ORP可以反映厭氧消化過(guò)程中細(xì)胞代謝活動(dòng)的氧化還原平衡和電子轉(zhuǎn)移。nZVI作為一種還原劑,可以保持厭氧系統(tǒng)中較低的ORP。產(chǎn)甲烷菌中的多種酶能適應(yīng)較低的ORP環(huán)境,防止酶被氧化而失去活性。由圖5(c)和(d)可以看出,含有nZVI的沼氣提純體系ORP顯著低于厭氧消化體系(P<0.05)。XU等的研究表明,體系ORP的降低是nZVI提高CH4產(chǎn)量的重要因素。此外,U2和U3系統(tǒng)的ORP低于U1,這可能也是S2和S3沼氣產(chǎn)量增加的原因之一。
富氫值(HE)表示液相中的氫含量(圖5(e,f))。與厭氧消化體系相比,沼氣提純體系內(nèi)HE由0.15~2.18mg·L-1(A1、A2、A3)增加到0.89~5.58mg·L-1 (U1、U2、U3)。這表明厭氧條件下nZVI的析氫將大幅度增加液相內(nèi)的H2含量,HE的增加可以提高H2的傳質(zhì)效率,促進(jìn)厭氧微生物將更多的H2和CO2轉(zhuǎn)化為CH4。此外,氣體循環(huán)促進(jìn)了更多H2和CO2在液相中的溶解,因而加速了微生物對(duì)H2和CO2的利用。
2.4 酶活的變化情況
表3為反應(yīng)結(jié)束后各體系酶活情況。A1、A2、A3中,堿性蛋白酶活性分別為17.67、62.66、67.48nmol·(min·g)-1(以VS計(jì)),乙酸激酶活性分別為74.91、79.07、91.55nmol·(min·g)-1,較初始污泥均有所提高。A1、A2、A3、U1、U2、U3中,脫氫酶的活性分別為10.40、15.16、29.29、30.34、31.23、31.10μg·(min·g)-1,較初始污泥顯著提高(P<0.05),且沼氣提純體系比厭氧發(fā)酵體系高。這是因?yàn)閚ZVI可以提供較低的ORP條件(圖5(d)),脫氫酶在該條件下能保持較高活性,有利于微生物高效利用H2和CO2,從而提高甲烷產(chǎn)率。輔酶F420是一種產(chǎn)甲烷菌酶,主要參與CH4的生成過(guò)程。U1、U2、U3中輔酶F420活性較原始污泥分別提高了84.85%、124.24%,127.73%,說(shuō)明氣體循環(huán)提高了輔酶F420的活性,尤其是生物提純體系。nZVI溶解產(chǎn)生的Fe2+也可以提高沼氣提純體系內(nèi)相關(guān)酶的活性,這對(duì)提高有機(jī)物的產(chǎn)CH4率具有促進(jìn)作用。
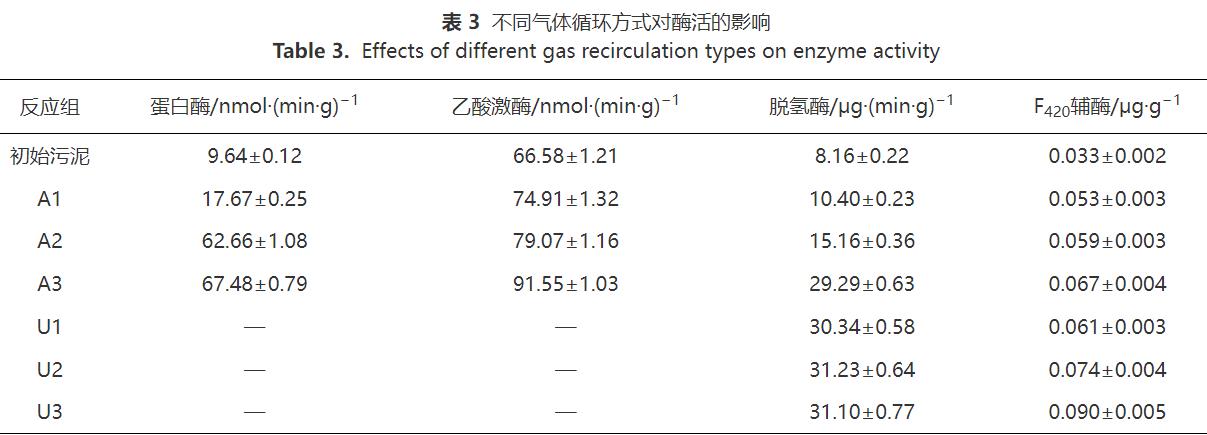
2.5 微生物群落變化分析
圖6(a)為反應(yīng)結(jié)束后各體系中細(xì)菌群落在門(mén)水平上的變化。初始污泥中豐度最大的是Chloroflexi(14.36%),其次是Actinobacteria(8.53%)。經(jīng)過(guò)厭氧消化和沼氣提純后,微生物結(jié)構(gòu)發(fā)生了較大變化。樣品中的優(yōu)勢(shì)菌主要由Chloroflexi、Firmicutes、Proteobacteria、Bacteroidetes、Actinobacteria組成。在厭氧消化體系內(nèi)(A1、A2、A3),Chloroflexi較初始污泥增加了5%~10%,Chloroflexi是厭氧消化的常見(jiàn)菌群,對(duì)餐廚廢水中的多糖和單糖均具有降解能力,并能夠產(chǎn)生乙酸為產(chǎn)甲烷菌提供底物。LI等的研究表明,Chloroflexi參與水解發(fā)酵過(guò)程,有利于促進(jìn)厭氧消化的進(jìn)程。Firmicutes提高了10.5%~14.4%,F(xiàn)irmicutes可以分泌多種胞外酶降解大分子底物,如蛋白質(zhì)和脂類(lèi)。因此,F(xiàn)irmicutes相對(duì)豐度的提高加速了難降解有機(jī)物的降解,為產(chǎn)甲烷菌提供營(yíng)養(yǎng)。Proteobacteria增加了2%左右,Proteobacteria的類(lèi)型比較復(fù)雜,其含有的多種類(lèi)型細(xì)菌不僅能利用葡萄糖、丙酸和丁酸等小分子化合物,而且會(huì)在酸化過(guò)程中產(chǎn)生乙酸為產(chǎn)甲烷菌提供底物。Bacteroidetes和Actinobacteria的相對(duì)豐度增加,Bacteroidetes和Actinobacteria是厭氧消化過(guò)程主要的產(chǎn)酸菌,對(duì)復(fù)雜的碳水化合物具有降解作用,其相對(duì)豐度的增加也會(huì)導(dǎo)致VFAs濃度的提高。這與圖4(b)的結(jié)果一致。沼氣提純體系內(nèi)(U1、U2、U3)由于沒(méi)有餐廚廢水作為底物,參與水解發(fā)酵過(guò)程的微生物沒(méi)有顯著變化,但與U1相比,U2和U3的5種優(yōu)勢(shì)菌相對(duì)豐度均有所增加。這說(shuō)明氣體循環(huán)可以提供H2和CO2,改善微生物群落結(jié)構(gòu)。
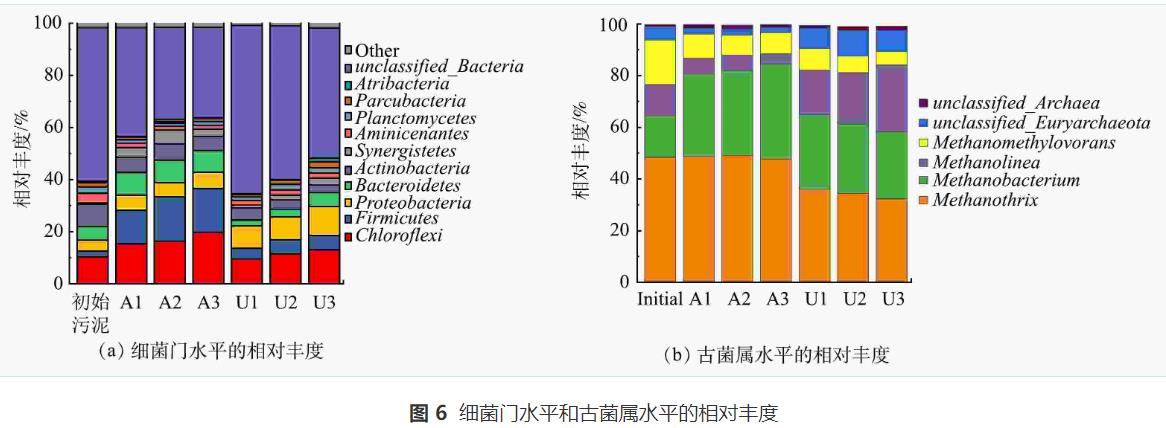
圖6(b)為反應(yīng)結(jié)束后各體系污泥中古菌群落在屬水平上的變化。優(yōu)勢(shì)菌是Methanothrix、Methanobacterium、Methanolinea、Methanomethylovorans。其中,Methanothrix屬于乙酸型產(chǎn)甲烷菌,Methanobacterium和Methanolinea屬于氫營(yíng)養(yǎng)型產(chǎn)甲烷菌,Methanomethylovorans屬于甲基型產(chǎn)甲烷菌。與原始接種污泥相比,Methanothrix在厭氧消化體系內(nèi)變化不大,但在沼氣提純體系降低了2%~6%,Methanomethylovorans在兩體系內(nèi)均有不同程度降低。然而,與初始污泥相比,Methanobacterium和Methanolinea由初始污泥的28%提高到38%~51%。此外,與厭氧消化體系(A1、A2、A3)相比,沼氣提純體系內(nèi)(U1、U2、U3) Methanobacterium和Methanolinea的相對(duì)豐度較大。這表明nZVI供氫的沼氣生物提純能促進(jìn)微生物向氫營(yíng)養(yǎng)型產(chǎn)甲烷菌演替。LI等認(rèn)為,Methanobacterium可以利用H2和CO2轉(zhuǎn)化為CH4。而與U1相比,U2和U3中2種氫營(yíng)養(yǎng)型產(chǎn)甲烷菌的相對(duì)豐度值最高。這表明氣體循環(huán)可以促進(jìn)氫營(yíng)養(yǎng)型產(chǎn)甲烷菌的富集,從而加快H2和CO2向CH4的轉(zhuǎn)化。
3、結(jié)論
1)對(duì)餐廚廢水厭氧消化產(chǎn)生的沼氣進(jìn)行基于nZVI供氫的異位生物提純,能有效提高CH4含量和產(chǎn)量。
2)單向式、往復(fù)式、循環(huán)式3種氣體循環(huán)方式的最終CH4含量分別為81.69%、88.12%、93.87%,CH4產(chǎn)量分別為57.65、85.12、157.18mL·g-1,其中循環(huán)式為最佳方式。
3)nZVI可降低沼氣提純體系內(nèi)ORP;nZVI供氫與氣體循環(huán)相結(jié)合,可以提高H2的氣-液傳質(zhì)效率;CO2再循環(huán)有利于維持體系酸堿平衡,優(yōu)化厭氧消化環(huán)境,在異位提純基礎(chǔ)上提高CH4含量和產(chǎn)量。
4)nZVI供氫的異位生物提純和氣體循環(huán)提高了脫氫酶和輔酶F420的相對(duì)活性,促進(jìn)了微生物向氫營(yíng)養(yǎng)型產(chǎn)甲烷菌的演替。其中,Methanobacterium和Methanolinea由初始污泥的28%提高到38%~51%。
廣東建樹(shù)環(huán)保科技有限公司是一家專(zhuān)業(yè)從事工業(yè)廢水處理、工業(yè)廢氣處理和環(huán)境修復(fù)的環(huán)保設(shè)備研發(fā)與銷(xiāo)售服務(wù)的企業(yè)。為工業(yè)企業(yè)和市政工程等項(xiàng)目提供工業(yè)廢水處理、工業(yè)廢氣處理、有機(jī)廢氣VOCs處理的一體化解決方案,從“工程設(shè)計(jì)”、“工程承包”、“設(shè)備采購(gòu)”、“安裝調(diào)試”、“耗材銷(xiāo)售”、“運(yùn)營(yíng)管理”、“環(huán)評(píng)辦理”等環(huán)節(jié)提供專(zhuān)業(yè)的差異化服務(wù),聯(lián)系電話(huà):135 5665 1700。





